(This post has resources and information for learning about Memory and Amnesia, for Physiological Psychology.)
Radio Lab:
6:30 Rat tests, 8:08 Chemical prevent memory in rats; Clive Waring 42:30
The Hippocampus and Patient H.M.(by Ted Ed)
H.M. - Nova special
9:22 What like for H.M. 10:40-11:52 Muscle memory star test
Morris Water Maze
Thursday, October 30, 2014
Friday, October 24, 2014
Periodic Table of Cookies
In honor of chemistry week,
Here's the Periodic Table of Cookies my club (Science Association of Women at UVU) did this spring to make money. Making that many cookies was insane. But fun. :)




Here's the Periodic Table of Cookies my club (Science Association of Women at UVU) did this spring to make money. Making that many cookies was insane. But fun. :)
 |
| Printed on sugar overlay, we cut each square out |
Tuesday, October 21, 2014
Footprints and trackways
How do paleontologists and anthropologists use footprints and trackways? Often, information is extrapolated from tracks and used to determine approximate heights and sizes of the organism that left the tracks. How can this be? Is there a correlation between foot size, leg length, stride length...and HEIGHT? Let's find out!
.JPG)

.JPG)

.JPG)


These formulae don't only work for dinosaurs. If you like Lego, you may like to know that using Alexander and Thulborn's formulae, a Lego minifig (the little people) could walk at 0.5 km/h and run at 2.2 km/h
(Lego info from http://www.csiro.au/helix/sciencemail/activities/dinospeed.html)
![Speed (in metres per second)=0.25 x (square root of g) x [stride length]^1.67 x [hip height]^-1.17](https://lh3.googleusercontent.com/blogger_img_proxy/AEn0k_uTBvbBbNFMWERgW-GHbJAkIIaz1fkAl6udC5csyojzIzbj70lMUvfKQshs8PMMpxh2GrMqguIpDnYZFBvyjOqT4IiDqSrrXTYpOV7-PaxULzyi_bXMuqj_uxUO4qfTlv_dr65jjczSEGVVwww=s0-d)
Where g is the acceleration from gravity. On Earth, g=9.8 m/s². Don't worry if you haven't come across maths like this before. What's important is that it's a way of estimating an animal's speed, without measuring it directly.
![Speed (in metres per second)=7.8 x ( 200/[number of steps] )^1.67 x [hip height]^-1.17](https://lh3.googleusercontent.com/blogger_img_proxy/AEn0k_v-sQhrkDzHNSHdtz83kiPYsa80igNCmNO0a9A6tsAqpuxlJi7SgI2zt8ojvv47czQznokMM-PWJEd5D8GoS96_PVpuUQD5K13u6obGzgEwxWq1cKcdLRPx5g7LqmUbZMNsmBCfQIGajEuNxwUM=s0-d) Your actual speed is just the distance divided by the time it took:
Your actual speed is just the distance divided by the time it took:

The basic measurement of a dinosaur footprint is its length, represented as FL. The ratio of footprint length and hip height (h) is different for different groups of dinosaurs, but generally the hip height of a bipedal dinosaur is roughly four times the footprint length. The speed can then be determined as relative speed, which is stride length (SL), divided by hip height (h). Generally speaking, if the SL/h <2.0, then the animal was walking; >2.9, the animal was running; and between 2.0 and 2.9, the animal was trotting.1 (Source: http://www.ucmp.berkeley.edu/education/dynamic/session3/sess3_act2.htm)
These formulae don't only work for dinosaurs. If you like Lego, you may like to know that using Alexander and Thulborn's formulae, a Lego minifig (the little people) could walk at 0.5 km/h and run at 2.2 km/h
 |
| Using the same equations that estimate dinosaurs' speeds, Lego figures could run at 2 km/h and walk at 0.5 |
![Speed (in metres per second)=0.25 x (square root of g) x [stride length]^1.67 x [hip height]^-1.17](http://www.csiro.au/helix/sciencemail/activities/images/DinoAlexander1.gif)
Where g is the acceleration from gravity. On Earth, g=9.8 m/s². Don't worry if you haven't come across maths like this before. What's important is that it's a way of estimating an animal's speed, without measuring it directly.
We can check Alexander's formula
by comparing it to our measurements. The stride length is just the
distance (100m) divided by the number of strides (half the number of
steps). In terms of your measurements, Alexander's formula becomes:
The basic measurement of a dinosaur footprint is its length, represented as FL. The ratio of footprint length and hip height (h) is different for different groups of dinosaurs, but generally the hip height of a bipedal dinosaur is roughly four times the footprint length. The speed can then be determined as relative speed, which is stride length (SL), divided by hip height (h). Generally speaking, if the SL/h <2.0, then the animal was walking; >2.9, the animal was running; and between 2.0 and 2.9, the animal was trotting.1 (Source: http://www.ucmp.berkeley.edu/education/dynamic/session3/sess3_act2.htm)
What can a single track tell us?
Not too much. We can infer something about the size of the animal and maybe something about the sediment. We may have a clue as to who made the track.What can a trackway (a series of tracks or footprints) tell you?
-
Who was there.
-
How many animals were there.
-
The sizes of the animals compared to one another.
-
How they were interacting - social activity, such as herds,
moving in families, etc.
-
How fast they were moving.
-
What the sediment was like, and therefore something about the
environment of the time.
Source of above text: http://www.ucmp.berkeley.edu/education/dynamic/session3/sess3_stories3.htm
Thursday, October 2, 2014
Leaf Anatomy
View of a shoot apical meristem with some leaf primordia. Additionally, I have labelled the 3 primary meristems you can differentiate here.

Here's a nicotine leaf to show the two kinds of meristems specific to the leaf. The leaf apical meristem becomes the midrib of the leaf, while the leaf marginal meristem is the blade of the leaf.

Epidermis
From a Sedum (stone crop)- you can see the "regular" epidermal cells (squiggly shaped) as well as the guard cells in various places surrounding stomata.

Take a closer look:

Also here is a sunken stomate as seen in a Pinus leaf. It is "sunken" because the guard cells are below the level of the epidermis which helps protect against dessication (drying out).
Mesophyll
Mesophyll just means middle of the leaf. It is the term for the ground tissue in the leaf. There are two types by shape: palisade and spongy.
You can see both of these well in a pine leaf:

We also term a leaf to be unifacial or bifacial based on the arrangement of the mesophyll. Bifacial is if there is spongy on one side, and palisade on the other. Unifacial is either the same type throughout, or it makes a kind of sandwich with the same kinds on either side and something different in the middle. You can see this type of unifacial in a Dianthus (carnation) leaf:

Pine anatomy
Some things we learned specifically with pine needles (I'm not sure if they also apply to some other plants, sorry) are the resin duct with epithelium, hypodermal sclerenchyma, and transfusion tissue.
Some of these are labeled:
The outermost layer surrounding the resin duct is the epithelium. Hypodermal sclerenchyma is below ("hypo") the outermost layer of cells or dermis. These have thick secondary cell walls which add to the strength of a pine needle, as you have probably tested yourself many times when you got poked with one. In this repeat picture you can see the hypodermal sclerenchyma with the thick red-stained walls, on either side of the sunken stomate.
Bundle Sheaths
Bundle sheaths are different in C3 plants and C4 plants.
Here, in a C3 grass (Poa), you see their regular bundle sheaths:

Closer. Just looks like a blank set of cells surrounding the vascular bundle.

But in a C4 plant, like this Zea corn, the sheaths have what is referred to as "Kranz anatomy". Kranz is German for wreath, and you can see they are rather leaf-like in the following examples. There are two orientation pictures, then it zooms in one a single bundle sheath so you can see the Kranz anatomy.
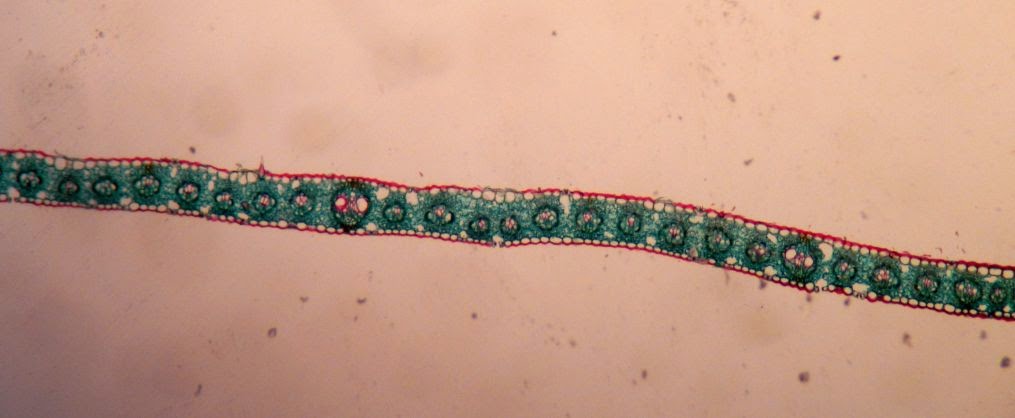



Leaf Abscission Zone
When a plant loses its leaves, it prepares for this by creating an abscission zone so the leaf can easily fall off without damaging any of the other tissue. Layers of cells secrete suberin (or is it subirin?) for protection (and are called suberized cells), while the next outermost layer of cells is pre-programmed to break easily, as it were. The weak layer is called the separation layer.

Whew, that's a lot of leaf anatomy! Stay curious.
P.S. There are no picture source references cause I took all these with my camera (through the microscope in Botany lab) and did the labeling myself.
Tuesday, September 9, 2014
Plant Cell Wall Lab
In this lab we explored various anatomical features of plant cell walls.
Mitosis & Cytokinesis
Not going to repost that, I did this last year and it is still beautiful: Mitosis post
Here's another great pic of a cell plate and phragmoplast on the edges I got today though:

Cytokinesis in plants is a rather involved process since a new middle lamella must be made, followed by primary wall and sometimes secondary wall.
Primary pit fields
This is a TERRIBLE picture taken of a picture shown to me on another camera. But you can kind of see the primary wall and the thinner parts that indicate the primary pit fields (pointer is on the thicker part of the primary cell wall).

More primary pit fields from a more 3D outside view of the cell (this is from #6 of the lab).

The cell walls are stained pink and you can see that there are lighter/ white spots on it. That's where the wall is thinner, so those are the primary pit fields.
Plasmodesmata
By definition, plasmodesmata are only in primary cell walls, and they are channels between adjacent cells, through which the cytoplasm of each cell is continuous. These occur more often in primary pit fields where the membrane is thinner, but they can happen anywhere in the primary wall.
 You can also see these in the tomato cells we looked at last week:
You can also see these in the tomato cells we looked at last week:

Intercellular Air Spaces
These get formed often at the edges of cells as they are dividing, and are important to gas exchange for the plant. (#6)

Secondary Cell Wall
Here you can see the distinction between the secondary wall and the compound middle lamella (which includes middle lamella and 2 adjacent primary walls).
 The lighter lines between all the cells are the compound middle lamella, the darker parts are secondary cell wall.
The lighter lines between all the cells are the compound middle lamella, the darker parts are secondary cell wall.
Pits
These are pear stone cells, and they have many long simple pits running through their secondary cell walls. I thought they look rather similar to plasmodesmata, but the distinction is plasmodesmata are ONLY in primary walls, and pits are ONLY in secondary walls. (Lab #9)

Bordered pits
Bordered pits from above look like little donuts. Here there are a whole bunch in some dense pine wood. These types of pits are common in water conducting cells, and they act to help prevent clogs from air bubbles. (Lab #11)


Stained microscope slides are pretty. That is all.
Stay curious.
Mitosis & Cytokinesis
Not going to repost that, I did this last year and it is still beautiful: Mitosis post
Here's another great pic of a cell plate and phragmoplast on the edges I got today though:
Cytokinesis in plants is a rather involved process since a new middle lamella must be made, followed by primary wall and sometimes secondary wall.
Primary pit fields
This is a TERRIBLE picture taken of a picture shown to me on another camera. But you can kind of see the primary wall and the thinner parts that indicate the primary pit fields (pointer is on the thicker part of the primary cell wall).
More primary pit fields from a more 3D outside view of the cell (this is from #6 of the lab).
The cell walls are stained pink and you can see that there are lighter/ white spots on it. That's where the wall is thinner, so those are the primary pit fields.
Plasmodesmata
By definition, plasmodesmata are only in primary cell walls, and they are channels between adjacent cells, through which the cytoplasm of each cell is continuous. These occur more often in primary pit fields where the membrane is thinner, but they can happen anywhere in the primary wall.
Intercellular Air Spaces
These get formed often at the edges of cells as they are dividing, and are important to gas exchange for the plant. (#6)
Secondary Cell Wall
Here you can see the distinction between the secondary wall and the compound middle lamella (which includes middle lamella and 2 adjacent primary walls).
 |
| Just a picture to orient- vascular bundle, and the bundle cap at the top. |
Pits
These are pear stone cells, and they have many long simple pits running through their secondary cell walls. I thought they look rather similar to plasmodesmata, but the distinction is plasmodesmata are ONLY in primary walls, and pits are ONLY in secondary walls. (Lab #9)
Bordered pits
Bordered pits from above look like little donuts. Here there are a whole bunch in some dense pine wood. These types of pits are common in water conducting cells, and they act to help prevent clogs from air bubbles. (Lab #11)
Stained microscope slides are pretty. That is all.
Stay curious.
Sunday, September 7, 2014
Plant Cell Wall Synthesis
Plants have some things animals don't, including a cell wall surrounding their cells.
Primary Cell Wall
Here's a diagram of the primary cell wall, along with the middle lamella that lies between adjacent plant cells with their respective primary cell walls. Also the regular-old plasma membrane lies internally to all that.
The middle lamella is made of pectins which are the perfect sticky thing to attach a primary wall made of cellulose microfibrils to! The primary wall also has some other stuff to hold it together in a nice meshy business.
Synthesis of Primary Cell Well
This is the coolest part...
When a cell splits and becomes two cells, a new cell wall must be built between them. I'll go into the details of the cytokinesis itself in another post, but after that is done, all that is there is is a middle lamella (again, made of pectins), and a plasma membrane on either side. How does the primary wall end up BETWEEN the plasma mebrane and the middle lamella?!?
Cellulose synthase, that's how. And it's brilliant. In the plasma membrane, there is a complex of proteins embedded that make cellulose. They look like little rosettes, like the ones depicted in blue, below:

The cellulose microfibrils get put together and come out of the external end of the rosettes (closer to the middle lamella). The long cellulose molecules that strengthen the primary wall adhere to the middle lamella, add some cross-linking stuff (pectin, glycans) and there you have it.
The cool part is those rosettes actually move through the plasma membrane, (like wading through mud) guided by microtubules which are on the internal side of the plasma membrane, leaving the trail of cellulose as it goes.
Here are some other diagrams of how this works.
In this one, the blue arrows indicate the direction the rosettes are "wading" through the plasma membrane, "walking" along the orange microtubules beneath.

This shows how the glucose subunits come in from the cytoplasm (purple circles) and are put together into the complex polymer of cellulose. Again, the arrow shows the rosette is moving to the left, leaving a trail of cellulose to the right.
And here we see the yellow plasma membrane cut away partly so we can see the rosettes that pass through and synthesize the microfibrils of cellulose. Each section of the rosette is an enzyme in its own right that puts together the long chains from glucose, which is then wound together into larger and larger units.
Here's what the structure of cellulose looks like broken down, so you can see it's a complex, tightly packed polymer.
The strands of cellulose are arranged pretty randomly in a primary cell wall. The cellulose synthases don't move very quickly, so the cellulose that is spit out goes around rather randomly, much like squeezing a bunch of toothpaste out of a tube- it goes every which way.
Secondary Cell Wall Synthesis
The secondary cell wall is lain down internally to the primary cell wall. It is thick and has 3 layers that are put down one at a time. Each layer has all its cellulose going parallel to each other. But the layers each have different directions/ orientations than one another, as seen in the bottom part of this diagram:
This provides a lot of extra strength, because it is protecting against compression, stretching, tension, etc. in all directions once you have all 3 layers put down. When the cellulose rosettes are laying down cellulose for a secondary cell wall layer, they move more quickly and in regular, straight lines.
That's all she (I) wrote. Stay curious!
 |
| Source of picture |
Primary Cell Wall
Here's a diagram of the primary cell wall, along with the middle lamella that lies between adjacent plant cells with their respective primary cell walls. Also the regular-old plasma membrane lies internally to all that.
 |
| Link to source |
Synthesis of Primary Cell Well
This is the coolest part...
When a cell splits and becomes two cells, a new cell wall must be built between them. I'll go into the details of the cytokinesis itself in another post, but after that is done, all that is there is is a middle lamella (again, made of pectins), and a plasma membrane on either side. How does the primary wall end up BETWEEN the plasma mebrane and the middle lamella?!?
Cellulose synthase, that's how. And it's brilliant. In the plasma membrane, there is a complex of proteins embedded that make cellulose. They look like little rosettes, like the ones depicted in blue, below:
The cool part is those rosettes actually move through the plasma membrane, (like wading through mud) guided by microtubules which are on the internal side of the plasma membrane, leaving the trail of cellulose as it goes.
Here are some other diagrams of how this works.
In this one, the blue arrows indicate the direction the rosettes are "wading" through the plasma membrane, "walking" along the orange microtubules beneath.
This shows how the glucose subunits come in from the cytoplasm (purple circles) and are put together into the complex polymer of cellulose. Again, the arrow shows the rosette is moving to the left, leaving a trail of cellulose to the right.
 |
| Source of image |
And here we see the yellow plasma membrane cut away partly so we can see the rosettes that pass through and synthesize the microfibrils of cellulose. Each section of the rosette is an enzyme in its own right that puts together the long chains from glucose, which is then wound together into larger and larger units.
 |
| Source of image |
Here's what the structure of cellulose looks like broken down, so you can see it's a complex, tightly packed polymer.
 |
| Source of image |
The strands of cellulose are arranged pretty randomly in a primary cell wall. The cellulose synthases don't move very quickly, so the cellulose that is spit out goes around rather randomly, much like squeezing a bunch of toothpaste out of a tube- it goes every which way.
Secondary Cell Wall Synthesis
The secondary cell wall is lain down internally to the primary cell wall. It is thick and has 3 layers that are put down one at a time. Each layer has all its cellulose going parallel to each other. But the layers each have different directions/ orientations than one another, as seen in the bottom part of this diagram:
 |
| Source of image |
This provides a lot of extra strength, because it is protecting against compression, stretching, tension, etc. in all directions once you have all 3 layers put down. When the cellulose rosettes are laying down cellulose for a secondary cell wall layer, they move more quickly and in regular, straight lines.
That's all she (I) wrote. Stay curious!
Subscribe to:
Posts (Atom)


{kind=link}
{kind=link}
{kind=link}